История исследований
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги , который это открытие опубликовал в 1675 году в своей работе Anatome plantarum . Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых . Прогресс в изучении наступил в XIX веке , и тогда же, в 1827 году , швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль , который открыл основной принцип открывания устьиц и Симон Швенденер , классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis ), Боб садовый (Vicia faba ), Кукуруза сахарная (Zea mays ).
Строение
Размеры устьица (длина) колеблются в пределах 0,01–0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum , 0,12 mm (120 µm) Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes) , которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis) . Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен . Воздух, содержащий диоксид углерода (углекислый газ) и кислород , проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями ) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный - сопровождающие клетки не отличаются от остальных клеток эпидермиса , тип весьма обычен для всех групп высших растений , за исключением хвойных ;
- диацитный - характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный - сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный - замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный - четыре сопровождающие клетки, характерен для однодольных ;
- энциклоцитный - сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный - несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный - замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный - замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный - замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный - устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный - такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
У двудольных распространённым является парацитный тип устьиц. Замыкающие клетки почковидной (бобовидной) формы - такими они видны с поверхности листа - несут хлоропласты , тонкие неутолщённые участки оболочки образуют выступы (носики) закрывающие устьичную щель.
Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном разрезе устьица. Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюдаются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, или внутренний, соединённый с крупным межклетником - подустьичной полостью.
У однодольных парацитное строение устьиц отмечено у злаковых . Замыкающие клетки имеют гантелевидную форму - сужены в средней части и расширены на обоих концах, при этом стенки расширенных участков очень тонкие, а в средней части замыкающих клеток сильно утолщены. Хлоропласты располагаются в пузыревидных окончаниях клеток.
Движение замыкающих клеток
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при неодинаковом водоснабжении в ночные часы, а иногда и днём тургор в замыкающих клетках понижается, и устьичная щель замыкается, снижая тем самым уровень транспирации . С повышением тургора устьица открываются. Считают, что главная роль в изменении тургора принадлежит ионам калия . Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и повышению тургорного давления в замыкающих клетках.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу . Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом - во время засухи.
У мхов (исключение антоцеротовые)настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Примечания
Литература
- Атлас по анатомии растений: Учеб. пособие для вузов / Бавтуто Г. А., Ерёмин В. М., Жигар М. П.. - Мн. : Ураждай, 2001. - 146 с. - (Учеб. и учеб. пособия для вузов). - ISBN 985-04-0317-9
- [[:en:{{{1}}}|]] Шаблон:Цитирование монографии
Сноски
Wikimedia Foundation . 2010 .
После прохождения устьица газ может диффундировать к поверхности клеток, находящихся внутри листа. Поверхность клеток защищена водной пленкой; внутри листа относительная влажность воздуха составляет 100%- Перенос газа к этим клеткам мезофила осуществляется с сопротивлением гмез- Сопротивление при переносе через граничный воздушный слой зависит от скорости ветра (она влияет на толщину слоя) и от диффузионных свойств газа .[ ...]
Вводные пояснения. Движение устьиц обусловлено особенностями их анатомического строения. Устьица состоят из двух замыкающих клеток полулунной или бобовидной формы, внутренние стенки которых утолщены, а наружные - тонкие. При насыщении замыкающих клеток водой наружные стенки сильно растягиваются, кривизна замыкающих клеток увеличивается и устьич-ная щель открывается. В случае потери воды замыкающие клетки выпрямляются и устьичиые щели закрываются. В основе динамики тургора замыкающих клеток лежит изменение их осмотического давления.[ ...]
Цель работы - исследование реакции устьиц регене рантов ex vitro в сравнении с интактными растениями. Объектом послужила твердая яровая пшеница сорта Харьковская 46. Использовали метод измерения усть-ичной проводимости (gs) с помощью порометра MK-Delta Т равновесного типа с непрерывным потоком воздуха. У регенерантов, выращиваемых в закрытых пробирках в условиях нормального освещения, замеряли gs в момент извлечения из пробирок. Затем регенеранты переносили в вегетационные сосуды с почвенной смесью на светоплощадку в условия слабого освещения. Сосуды в первые 1-3 сут накрывали стеклянными колпаками для создания аналогичной пробиркам высокой ОВВ (99%), т.е. в условия пониженного транспирационного запроса. Затем, приоткрывая сосуды, ОВВ постепенно уменьшали. Через 10 сут стеклянные колпаки снимали, и регенерантам создавали условия нормального освещения. В течение 1-15 сут ежедневно замеряли gs. Установлено, что gs регенерантов, находящихся в пробирках, в 2 раза выше gs интактных растений, что, возможно, связано с высокой ОВВ в закрытых пробирках. Но все же после процесса адаптации на 7-9-е сут устьица регенерантов были открыты шире на 20%, чем устьица интактных растений. При нормальном освещении gs регенерантов повышался, что, по-видимому, связано с действием света на устьица. Однако gs регенерантов был на 20% ниже, чем у интактных растений, т.е. устьица регенерантов не были способны открыться достаточно широко.[ ...]
Небольшие, дозы 302 вызывают открытие устьица листа, при высоких дозах происходит их закрытие . В соответствующих условиях увеличение времени пребывания устьиц в открытом состоянии приводит к водному стрессу, но связь такого физиологического явления с метаболизмом листа абсолютна неясна. Влияние Э02 на метаболизм листа в большинстве случаев изучалось в в доиллимольных концентрациях сульфитных растворов. Связь концентрации БСЬ в газовой фазе с его равновесной концентрацией, установленная с учетом возможных реакций в растворе (исключая окисление), представлена на рис. УП-5. При концентрации БОг в газовой фазе равной 100 мкг/м3 (0,3 млн-1), равновесная концентрация соединений Э(1У) в растворе составляет 36 ммоль/дм3. Эта величина в действительности могла быть выше, если можно было бы оценить и учесть буферную способность мезофильной воды.[ ...]
Как известно, для газообмена и транспирации растению служат устьица - особые клетки, расположенные в эпидермисе, их иногда образно называют «форточками» растений. Так вот количество устьиц у суккулентов обычно уменьшено, кроме того, устьица иногда располагаются не на поверхности, а в глубине ткани листа.[ ...]
Травы или вторичные древовидные формы. Устьица ано-моцитные или с побочными клетками, обычно с 2 побочными клетками (нарацитные). Сосуды только в корнях или во всех вегетативных органах, очень редко отсутствуют. Цветки обоеполые или редко однополые. Околоцветник хорошо развит и состоит из сходных между собой (обычно лепестковидных) или ясно различающихся чашелистиков и лепестков, или же околоцветник редуцирован. Зрелая пыльца обычно 2-клеточная, реже 3-клеточная. Гинецей, как правило, ценокарпный, редко (у примитивных триурисовых и некоторых примитивных лилейных) более или менее апокарпный. Семена обычно е обильным эндоспермом, по у порядка имбирных с периспермом и остатком эндосперма или только с периспермом.[ ...]
Большее нагревание листьев, чем окружающего воздуха, способствует испарению. Устьица - отверстия, через которые проходит воздух и пары воды, у этого растения находятся на нижней стороне листа.[ ...]
У S. falcispora пикниды оливково-бурые, приплюснуто-шаровидные, 50-100 мкм в диаметре, с маленьким круглым устьицем. Пикноспоры бесцветные или желтые, с 5-7 перегородками, прямые или серповидно согнутые, размером 30-43X3-4 мкм. У S. secalis пикниды расположены рядами, диаметр их до 90 мкм. У S. nodorum диаметр пикниды варьирует от 70 до 210 мкм, располагаются они обычно вдоль жилок листа, имеют слабо развитые устьица; иикноспоры узкоцилиндрические, бесцветные, с 3 перегородками, размером 15-32X2-4 мкм. Зимуют грибы в виде грибницы и иикнид с пикноспорами на всходах озимой ржи и остатках пораженных растений у поверхности почвы. При наличии капельной влаги и температуре от 9 до 22°С пикноспоры прорастают. Заболевание приводит к преждевременному усыханию листьев, недоразвитости колоса и ощутимому недобору зерна (до 15%). Устойчивых к септориозу сортов ржи нет.[ ...]
Возбудитель болезни - бактерии Xanthomonas campestris pv. vesicatoria Dye (==Xant. vesicatoria Dowson). В растение проникают через устьица и места ранения, а распространяются по межклеточным пространствам мезофилла и полисадной ткани листа. Продолжительность инкубационного периода болезни при заражении листьев 3-6 дней, а при заражении плодов - 5-6 дней.[ ...]
Многолетние травы или кустарнички, иногда лианы с очередными цельными листьями, лишенными прилистников. Устьица разных типов. Членики сосудов с простой перфорацией. Цветки в верхоцветных соцветиях или в кистях, обоеполые, 5-членные, с двойным околоцветником. Чашечка сростнолистная, большей частью сухопленчатая. Венчик обычно явственно сростнолепестный. Тычинок 5, супротивных лепесткам и более пли менее приросших к трубке венчика. Пыльцевые зерна 3-5-бо-роздные или реже (¡-рассеянно-бороздные. Гинецей цеиокарпный (лизикарпный), из 5 плодолистиков, со свободными или более или менее сросшимися столбиками; завязь верхняя, с одним базальным семязачатком на длинной, почти окружающей его семяножке. Характерно наличие-обтуратора (пробочки), отходящего от верхней части завязи н врастающего в микропиле (он содействует прохождению пыльцевой трубки). Плод сухой, нераскрывающийся или редко раскрывающийся створками снизу вверх. Семена с большим прямым зародышем и обычно с ондоспормом, по без перисперма. Происхождение, вероятно, от примитивных представителей порядка гвоздичных, скорее всего от предков, близких к современным портулаковым и базелловым.[ ...]
Число устьвчных отверстий колеблется в зависимости от вида растения от 1 до 60 тыс. на 1 см2 листа. Большая часть устьиц расположена на нижней стороне листа.[ ...]
Деревья, кустарники или травы. Листья цельные или различным образом расчлененные. Устьица разных типов, большей частью без побочных клеток. Сосуды всегда имеются; членики сосудов с лестничной или простой перфорацией. Цветки обоеполые или однополые, с двойным околоцветником или реже безлепестные; у более примитивных семейств околоцветник часто спиральный или спироциклический. Андроцей, когда он состоит из многих тычинок, развивается л центробежной последовательности. Зрелая пыльца 2-клеточная или реже 3-клеточная. Гинецей апокарпный или чаще цепокаршшй. Семена, как правило, с эндоспермом.[ ...]
Источником инфекции являются зараженные семена и неперегнившие растительные остатки. Заражение происходит через устьица и ранки. Инкубационный период болезни 5 - 10 дней. При сильном ее развитии недобор урожая плодов может составлять 40 % и более.[ ...]
Зооспоры могут жить и двигаться только в воде. Попав на зеленые части растения, они быстро передвигаются к открытым устьицам, затем втягивают в себя жгутики, облекаются тонкой оболочкой и прорастают, давая нитевидный росток, внедряющийся в ткань растения через устьице. Росток разрастается в грибницу, располагающуюся по межклеткам. В клетки растения-хозяина заходят гаустории, с помощью которых гриб извлекает питательные вещества. По мере роста грибницы в начале отмирания клеток растения формируются органы размножения. Летом при соответствующей влажности гриб обычно образует бесполое спороношение - зооспораигиеносцы с зооспорангиями, которые выходят через устьица и образуют белый налет.[ ...]
Однако это не единственное объяснение. В последнее время японский исследователь М. Фужино показал, что замыкающие клетки устьиц в открытом состоянии содержат значительно больше калия па свету но сравнению с темнотой. Прибавление АТФ к эпидермису, плавающему на растворе КО, увеличивает скорость открытия устьиц иа свету. На основании этих данных можно считать, что осмотическое давление замыкающих клеток устьиц воврастает благодаря усиленному поступлению калия, регулируемому АТФ. Причем для этого может быть использована АТФ, образованная в процессе фотосипте-тического фосфорилирования, происходящего на свету (с. 126). Вов-иожно, что обе эти теории являются верными. Не вызывает сомнений необходимость АТФ для осуществления движения устьичпых клеток. Исследования советского ученого С. А. Кибрика показали возрастание содержания АТФ в замыкающих клетках устьиц в процессе их открывания. Указанные процессы представлены в виде схемы.[ ...]
Винтеровые - деревья и кустарники с цельными, перистонервными, прозрачно-точечными, кожистыми листьями, лишенными прилистников. Устьица на листьях с 2 побочными клетками, т. е. наиболее примитивного типа. Замечательной особенностью вииторовых является отсутствие сосудов во всех органах растения. Водопроводящие элементы ксилемы состоят из очень длинных и толстостенных трахеид.[ ...]
У многих родов стенка спорогона состоит из нескольких слоев клеток (например, у анто-цероса). В ее наружном плотном слое расположены устьица, образованные двумя замыкающими клетками. Имеются и редуцированные формы спорогонов, у которых нет устьиц (нототилас). Центральная часть коробочки спорогона состоит из бесплодной колонки, но последняя обычно отсутствует у того же нототиласа. Между колонкой и стенкой коробочки развиваются тетрады спор и бесплодные нити- элатеры. Существует мнение, что колонка выполняет механическую функцию и служит также для проведения воды и питательных веществ. Образована она продольно вытянутыми клетками с узкими просветами.[ ...]
В зоне пятна из каждого устьица выходят обычно 4-5 зооспорангиеиосцев, но их число может достигать 20.[ ...]
Передвижение воды и питательных веществ вверх по ксилеме у высших растений частично связано с транспирацией, т. е. испарением влаги листьями через многочисленные устьица. По мере потери воды клетками недостаток диффузионного давления притягивает воду из элементов ксилемы, которые образуют крупные многочисленные сплошные трубки (сосуды) от корней до листьев. Таким образом, натяжение передается через весь столб к клеткам корня и приводит к усилению поглощения воды. Скорость транспирации зависит от степени раскрытия устьиц и от таких окружающих факторов, как температура и влажность воздуха, которые влияют на физическую скорость испарения воды. Замыкание и размыкание устьиц является механическим процессом, регулируемым тургором замыкающих клеток (см. рис. 27).[ ...]
В период вегетации растений распространяется конидиями, образующимися на конидиеносцах. Конидиеносцы пяти-шести-кратно вильчато-разветвленные, выступают на поверхность листьев из устьиц одиночно или по 2-3. Конидии светло-фиолето-вые, яйцевидной формы, размером 20-28Х 17-23 мкм (рис. 32).[ ...]
В лесном хозяйстве снижается прирост древесины, отмирают менее устойчивые древесные породы, а в отдельных случаях наблюдается гибель леса на целых участках. Вредные газы, проникая через устьица листа, нарушают процессы фотосинтеза и дыхания. Так, концентрация сернистого ангидрида в воздухе в соотношении 1:1000 000 при повторных действиях в течение двух месяцев вызывает повреждение растений. Более высокие концентрации могут приводить к полной потере листьев растениями.[ ...]
Перитеции - плодовые тела закрытого типа, имеющие вид замкнутого кувшина округлой или овальной формы с маленьким отверстием в верхней части (рис. 307). Через это отверстие, которое носит название устьица, происходит выбрасывание зрелых спор. Обычно перитеции почти незаметны на поверхности лишайника: при внимательном рассмотрении можно обнаружить лишь черные точки - устьица перитециев, в то время как само плодовое тело целиком погружено в слоевище. Реже перитеции выступают своими верхушками (рис. 307,1) или целиком сидят на слоевище. Стенки этого кувшинообразного плодового тела состоят из нескольких слоев (рис. 307,2). Внутреннюю полость перитеция, где образуются сумки со спорами, окружает эксципул. Это темно- или светлоокрашенная оболочка, состоящая из нескольких слоев гиф, поделенных на удлиненные клетки. Нередко эксципул снаружи покрыт еще одной оболочкой - п о к р ы-вальцем. Покрывальце, как правило, бывает темноокрашенным или черным, может охватывать эксципул со всех сторон или покрывать его лишь до половины; иногда оно развито лишь у устьица, а нередко и вовсе отсутствует. Внутри перитеция развиваются сумки со спорами, которые в нижней и боковой частях плодового тела образуют вместе с парафизами гимениальный слой (рис. 307, 26). Гимениальный слой развивается на гипотец и и - узком зернистом слое, прилегающем к внутренней стенке перитеция. Характерно, что у некоторых лишайников в перитециях защитные нити - парафизы - вовсе не образуются или очень рано расплываются в слизь. Это связано с тем, что сумки со спорами надежно защищены самими стенками перитеция и надобность в парафизах исчезает. Особые защитные нитевидные гифы в перитециях развиваются около устьица (рис. 307, 2г). Они называются перифизами и служат для защиты всего внутреннего ядра перитеция от воздействий внешней среды.[ ...]
С другой стороны, работы с одним мутантом томата («флакка» - завядающий) позволили сделать интересные наблюдения, указывающие на регуляторную роль АБК в процессе транспирации. У таких мутантов даже при нехватке воды устьица остаются открытыми, так что растения постоянно находятся в состоянии завяда-ния. После опрыскивания листьев раствором АБК симптомы завядания исчезали. Устьица закрывались, водный баланс выравнивался, и растения вновь становились тургесцентными.[ ...]
Озон действует на ту часть листа, которая находится прямо под кожицей. Появляются характерные пятна, сливающиеся в своеобразный узор. Некоторые виды содержащихся в смоге газов действуют на нижнюю поверхность листа, проходя сквозь устьица и также вызывая появление полосок омертвевшей ткани. Все эти повреждения листвы легко заметить. Но во всех случаях, кроме полной гибели, никак нельзя сказать, насколько сильно вредные газы ухудшают общую продуктивность растений. О тех скрытых ранах и рубцах, которые остаются на растении после битвы с грязным воздухом, известно очень мало.[ ...]
В экзотеции нижней половины урночки и шейки находятся устьица. Различают устьица поверхностные, расположенные на уровне экзотеция, и погруженные, расположенные ниже его уровня. Устьица обычной формы имеют щель и состоят из двух симметричных почковидных замыкающих клеток. Устьица без щели присущи сфагновым.[ ...]
В среднем сухое вещество растений содержит около 45% углерода и 42% кислорода. Источником углерода и кислорода для синтеза органических веществ растения является воздушное питание. Углекислый газ проникает вместе с воздухом в листья через «устьица», густо усеивающие листовую пластинку. Одновременно через устьица идет испарение воды. Суммарная поверхность листьев превосходит (в 20-70 и больше раз) площадь почвы, занимаемую растением, что создает хорошие условия для поглощения С02 и энергии солнечных лучей зелеными листьями. Этот цвет зависит от хлорофилла, космическую роль которого убедительно раскрыл К. А. Тимирязев, ибо без хлорофилла растения не могли бы улавливать энергию солнечных лучей, а следовательно, и запасать ее в форме потенциальной энергии урожая.[ ...]
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ - гуттация. Выделение жидкости идет через специальные водные устьица - гидатоды. Выделяющаяся жидкость - гутта. Таким образом, процесс гуттации является результатом одностороннего, тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.[ ...]
Однолетнее растение мятлик луговой (Роа annua) является одним из наиболее чувствительных к смогу, и его повсеместное распространение делает его весьма удобным индикатором смога. Поражения ограничены теми «летками, которые находятся в стадии максимального расширения. Они возникают на верхушке наиболее молодых листьев и постепенно переходят и на старые листья. Начальное и наиболее тяжелое поражение возникает в окружении подустьичных камер. За дезинтеграцией хлоропластов следует плазмолиз и, наконец, полная дегидратация пораженных клеток, что приводит к мумификации мезофильной ткани в пораженных участках.[ ...]
У низших групп высших растений устьич-ный аппарат имеет очень примитивное строение, но в процессе возрастающего приспособления к воздушной среде он постепенно эволюционировал. Увеличивается неравномерность утолщения стенки замыкающих клеток и совершенствуется механизм регуляции устьица. У высших групп возникают так пазываемые побочные клетки, представляющие собой соседние с замыкающими клетками специализированные эпидермальные клетки, отличающиеся морфологически от соседних клеток эпидермы. Функционально, а в ряде случаев и по образованию в процессе онтогенеза (см. пиже), они тесно связаны с замыкающими клетками. Устьице вместе с побочными клетками (когда они присутствуют) называют у с т ь и ч-ным аппаратом или устьичным комплексом.[ ...]
Почки дафнифиллума снабжены черепитчатыми чешуями. Листья цельные, длинпоче-решковые, перистонервные, обычно кожистые, темно-зеленые, с восковым налетом, снизу часто голубовато-сизые, иногда с сосочками. Листья сохраняются немногим больше года и опадают вскоре после распускания молодых листьев. В листьях и стеблях имеются идиобласты, содержащие кристаллы. Перфорация члеников сосудов лестничная, с 20-30 и более тонких перекладин.[ ...]
Листья тиссовых ланцетовидные или линейные (даже узколинейные), с низбегающим основанием, иногда на коротких (1-2 мм) черешках. На верхней стороне листа посредине тянется продольное углубление, на дне которого чуть выступает средняя жилка, снизу же между хорошо выраженной средней жилкой и зелеными, лишенными устьиц полосками по краям листа проходят две светлые устьичные полоски, то узкие, то более широкие.[ ...]
Болезнь вызывают бактерии Pseudomonas syringae pv. coro-nafaciens Young, et al. (-Ps. coronafaciens Stevens). Эти палочковидные, одиночные или собранные в короткие цепочки грам-отрицательные бактерии имеют один или несколько полярных жгутиков. Развиваются при температуре от 0 до 31 °С (оптимум 24-25 °С), а погибают при 47-48 °С. В растения проникают через устьица и механические повреждения. В период вегетации растений возбудитель болезни распространяется при помощи ветра и с каплями дождя. Вредоносность болезни в основном заключается в уменьшении ассимиляционной поверхности растений, что приводит к недобору урожая. Иногда снижается всхожесть семян (на 2-5 %).[ ...]
Листья у акмопиле, как и у предыдущего рода, диморфные. При этом линейные фотосинтезирующие листья, так же как у фалькатифолиумов тиссовидного и серповидного, сплющенные с боков. Однако имеются и отличия. Так, палисадная паренхима здесь развивается лишь с одной стороны листа, ставшей (в ходе описанного выше превращения) верхней. Соответственно устьица можно видеть преимущественно на его поверхности, ставшей нижней (рис. 221).[ ...]
Зимуют яйца преимущественно на концах веток и плодовых образований. Во время распускания почек отрождаются личинки и первые два-три дня питаются на поверхности, а затем проникают внутрь почек. Личинки высасывают листья и бутоны, выделяют сахаристые экскременты в виде капелек росы, так называемую «медвяную росу», которая склеивает листья, бутоны и закупоривает устьица на листьях. На этих клейких выделениях нередко поселяются грибы, покрывающие сажистым налетом листья. Вследствие.высасывания медяницей клеточного сока листья недоразвиваются, бутоны не раскрываются, завязи осыпаются. После цветения яблони появляются взрослые насекомые, которые первые дни держатся скученно на нижней стороне листьев, а затем перелетают на другие деревья и на травянистую растительность. В начале августа медяница возвращается на яблоню и откладывает яйца.[ ...]
Проводящая система стебля - редуцированная сифоностела, которая часто принимает форму протостелы. Большая часть стебля занята корой, лишенной межклеточных пространств. Строение листа азоллы указывает на высокую специализацию. Каждый лист состоит из двух лопастей, или сегментов. Верхний сегмент, выступающий над водой, зеленый, из нескольких слоев клеток в толщину, с устьицами на обеих сторонах. Нижний сегмент погружен в воду. Он служит, как предполагают, для всасывания воды. На некоторых нижних сегментах развиваются сорусы.[ ...]
Освещенность отражается и на различии в анатомическом строении листьев световых и теневых растений (рис. 43). Листья световых растений имеют хорошо выраженную палисадную ткань, а если растение живет на почве, отражающей много света (мел, известняк), то палисадная ткапь может быть развита с обеих сторон листа. У световых растений эпидермис состоит из относительно мелких тонкостенных клеток, количество устьиц относительно велико. Скиофиты представляют противоположность гелиофитам и по этим признакам. Однако подобные различия связаны не столько с влиянием света, сколько с тем, что, живя на свету, скиофиты сильно нагреваются и это очень отражается на их водном режиме - световые растения часто являются в то же время растениями засушливых мест - ксерофитами.[ ...]
О механизме действия масел. При исследовании механизма токсического действия гербицидных масел и причин их избирательности было установлено, что мор-фолого-анатомические особенности растений не имеют большого значения для избирательного действия масел. Они легко и быстро смачивают все растения, как чувствительные, так и устойчивые. Они также легко проникают в ткани всех растений, главным образом через устьица и частично через кутикулу, заполняя межклеточные пространства. Таким образом, контакт масла с клетками обеспечивается в равной мере у обеих групп растений.[ ...]
Возбудитель картофельной болезни - гриб Phytophthora infestans (рис. 30, табл. 6) - развивает внутри листьев картофеля межклеточную грибницу с гаусториями. Питаясь тканями листа, он вызывает образование темных пятен, которые во влажную погоду чернеют и загнивают. При сильном поражении отмирает весь лист (табл. 6). Сначала погибают нижние листья, а затем болезнь захватывает и все растение. Попадая в капли воды на поверхности листа картофеля, спорангии прорастают шестью - восемью зооспорами, которые после периода движения округляются, покрываются оболочкой и прорастают ростковой трубкой (рис. 30, 1-4). Росток через устьице проникает в ткань листа (рис. 30,5). При благоприятных условиях (прохладная дождливая погода или сильные росы) время от заражения до образования нового спороно-шения составляет всего 3-4 дня. Так как вокруг одного пятна образуются тысячи зооспорангиев, можно понять, насколько быстро распространяется болезнь в посевах картофеля.[ ...]
Критерием солеустойчивости растений служит степень снижения продуктивности при засолении по сравнению с продуктивностью на нормальном фоне. Определяют солеустойчивость прямыми, а также менее трудоемкими косвенными методами. На засоленных почвах обычно снижается всхожесть семян, поэтому оценку солеустойчивости выполняют по показателям прорастания семян (энергия прорастания, процент всхожести, скорость прорастания). Из косвенных лабораторных методов наиболее известны: плазмолитический; определения скорости раскрытия устьиц в растворах солей; степени и скорости «выцветания» хлорофилла, количества альбуминов, проницаемости протоплазмы; биохемилюминесценция и др.[ ...]
Для комбретовых характерно наличие жолб-зок на черешке листа или на пластинке у основания, по сторонам главной жилки или у края. У некоторых видов вся пластинка усеяна мелкими желёзками, отчего лист выглядит точечно-прозрачным. Не менее характерно для комбретовых опушение из простых или многоклеточных волосков, выделяющих оксалаты кальция. Строение волосков разнообразно и является систематическим признаком (рис. 112). Наличие многоклеточных волосков в виде щитовидных чешуек на ножке, присущее видам комбретума и тилоа (ТЫ1оа), позволяет определять даже фрагменты их ископаемых остатков. Членики сосудов с простой перфорацией.[ ...]
Уредопустулы - продолговатые или округлые подушечки ярко-оранжевого цвета. Уредоспоры в них вначале прикрыты эпидермисом, который затем разрывается и споры имеют возможность рассеиваться. Уредоспоры ярко-желтые, шаровидные, покрытые шипиками. По краям в уредопустулах находятся булавовидные нити - парафизы. Споры, попавшие на новые растения, прорастают в каплях росы и посредством ростковой трубки внедряются через устьица в ткани растения-хозяина. Проявление симптомов заболевания наблюдается приблизительно через две недели после заражения.[ ...]
Обитание растений в водной среде, помимо перечисленных выше особенностей, накладывает отпечаток и на другие стороны жизнедеятельности, особенно на водный режим у растений, в прямом смысле окруженных водой. У таких растений транспирации нет, а следовательно, и нет «верхнего двигателя», поддерживающего ток воды в растении. И вместе с тем ток, доставляющий к тканям питательные вещества, существует (правда, значительно более слабый, чем у сухопутных растений) с ясно выраженной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению (у прикрепленных видов) и деятельности специальных клеток, выделяющих воду, - водяных устьиц или гидатод.[ ...]
Риниофитовую гипотезу отнюдь нельзя считать доказанной, и ее принимают далеко не все ботаники. Но если она в конце концов подтвердится полностью, то спорофит моховидных можно будет истолковать как конечный результат редукции дихотомически разветвленного спорофита предковых форм. С этой точки зрения спорогон будет соответствовать одной конечной веточке риниофитов. Если моховидные действительно произошли от риниофитов, то редукция спорофита была, вероятно, результатом приспособления к избыточной влажности. Именно этим можно было бы объяснить также характерное-для моховидных крайнее упрощение и часто-даже полное исчезновение проводящей системы. А то, что проводящая система должна была быть относительно хорошо развитой и нормально функционировать, доказывается частым наличием у моховидных устьиц, обычно рудиментарных. Ведь устьице коррелятивно связано с проводящей системой. Трудно себе представить, чтобы такое сложное приспособление могло возникнуть у спорофитов, не обладающих хорошо развитой проводящей системой и не ведущих самостоятельную жизпь вполне автотрофного поколения. Поэтому наличие устьиц является веским доводом в пользу происхождения моховидных от растений с хорошо развитым спорофитом. Такими растениями могли быть, скорее всего, риниофиты.[ ...]
Насосная функция растений может ослабляться или усиливаться в зависимости от условий внешней среды. Открытием чрезвычайной важности было установление анти-транспирационного эффекта С02. Этому явлению, которое несомненно должно иметь как положительные, так и серьезные теневые стороны, пока уделяется слишком мало внимания. Опыты с пшеницей и кукурузой показали, что при повышении концентрации С02 с 300 до 600 частей на млн. транспирация у этих культур снижалась соответственно на 5 и 20%. Оказалось также, что по увеличению эффективности использования воды в условиях дополнительной подкормки углекислотой кукуруза превосходит хлопчатник. С помощью специальной аппаратуры была установлена причина указанного явления. Оказалось, что рост концентрации С02 в воздухе вызывает снижение устьич-ной проводимости и повышает эффективность использования воды у всех растений, подвергающихся опытам (рис.2). Устьица - это маленькие отверстия на поверхности листьев, обычно около 10 мкм длиной и от 2 до 7 мкм шириной. Через них растеши осуществляют газообмен с атмосферой.
Лист — чрезвычайно важный орган растения. Лист — часть побега. Основными функциями его являются фотосинтез и транспирация. Лист характеризуется высокой морфологической пластичностью, разнообразием форм и большими приспособительными возможностями. Основание листа может расширяться в виде косых листовидных образований — прилистников с каждой стороны листа. В некоторых случаях они настолько велики, что играют определённую роль в фотосинтезе. Прилистники бываю свободными или приросшими к черешку, они могут смещаться на внутреннюю сторону листа и тогда их называют пазушными. Основания листьев могут быть превращены во влагалище, окружающее стебель и препятствующие его сгибанию.
Внешнее строение листа
Листовые пластинки различаются по размерам: от нескольких миллиметров до 10-15 метров и даже 20 (у пальм). Продолжительность жизни листьев не превышает нескольких месяцев, у некоторых — от 1,5 до 15 лет. Размер и форма листьев являются наследственными признаками.
Части листа
Лист — боковой вегетативный орган, растущий от стебля, имеющий двустороннюю симметрию и зону роста при основании. Лист обычно состоит из листовой пластинки, черешка (исключением являются сидячие листья); для ряда семейств характерны прилистники. Листья бываю простые, имеющие одну листовую пластинку, и сложные — с несколькими листовыми пластинками (листочками).
Листовая пластинка — расширенная, обычно плоская часть листа, выполняющая функции фотосинтеза, газообмена, транспирации и у некоторых видов — вегетативного размножения.
Основание листа (листовая подушка) — часть листа, соединяющая его со стеблем. Здесь находится образовательная ткань, дающая рост листовой пластинке и черешку.
Прилистники — парные листовидные образования в основании листа. Они могут опадать при развёртывании листа или сохраняться. Защищают пазушные боковые почки и вставочную образовательную ткань листа.
Черешок — суженная часть листа, соединяющая своим основанием листовую пластинку со стеблем. Он выполняет важнейшие функции: ориентирует лист по отношению к свету, является местом расположения вставочной образовательной ткани, за счёт которой растёт лист. Кроме этого, он имеет механическое значение для ослабления ударов по листовой пластинке от дождя, града, ветра и пр.
Простые и сложные листья
Лист может иметь одну (простой), несколько или множество листовых пластинок. Если последние снабжены сочленениями, то такой лист называется сложным. Благодаря сочленениям на общем черешке листа листочки сложных листьев опадают поодиночке. Однако у некоторых растений сложные листья могут опадать и целиком.
По форме цельные листья, различают как лопастные, раздельные и рассечённые.
Лопастным называю лист, у которого вырезы по краям пластинки доходят до одной четверти его ширины, а при большем углублении, если вырезы достигают более четверти ширины пластинки, лист называется раздельным. Лопасти раздельного листа называют долями.
Рассечённым называют лист, у которого вырезы по краям пластинки доходят почти до средней жилки, образуя сегменты пластинки. Раздельные и рассечённые листья могут быть пальчатые и перистые, дважды пальчатые и дважды перистые и т.д. соответственно этому различают пальчато-раздельный лист, перисторассечённый лист; непарно-перисторассечённый лист у картофеля. Он состоит из конечной доли, нескольких пар боковых долек, между которыми располагаются ещё меньшие дольки.
Если пластинка удлинённая, а доли или сегменты её треугольные, лист называют струговидным (одуванчик); если боковые доли неравновеликие, к основанию уменьшаются, а конечная доля крупная и округлая, получается лировидный лист (редька).

Что касается сложных листьев, то среди них различают тройчатосложные, пальчатосложные и перистосложные листья. Если сложный лист состоит из трёх листочков, он называется тройчатосложным, или тройчатым (клён). Если черешочки листочков прикрепляются к главному черешку как бы в одной точке, а самые листочки расходятся радиально, лист называется пальчатосложным (люпин). Если на главном черешке боковые листочки расположены с обеих сторон по длине черешка, лист называется перистосложным.
Если такой лист заканчивается наверху непарным одиночным листочком, получается, непарноперистый лист. Если же конечного нет, лист называется парноперистым.
Если каждый листочек перистосложного листа, в свою очередь, является сложным, то получается дважды перистосложный лист.
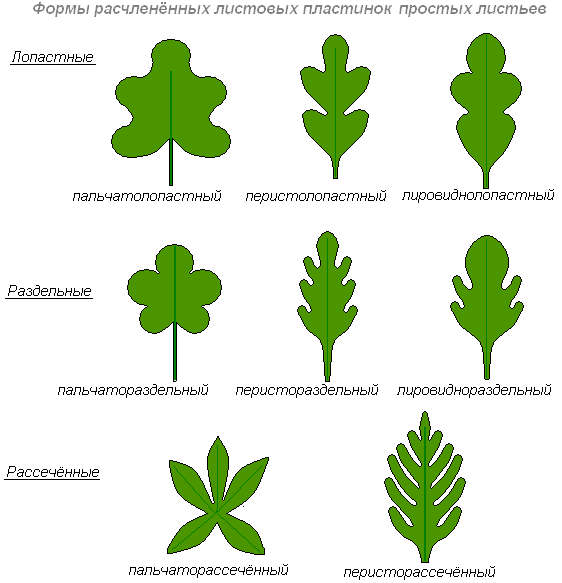
Формы цельных листовых пластинок
Сложным листом называют такой, на черешке которого имеется несколько листовых пластинок. Они крепятся к главному черешку своими собственными черешками, нередко самостоятельно, поодиночке, опадают, и называются листочками.
Формы листовых пластинок различных растений отличаются по очертанию, степени расчленённости, форме основания и верхушки. Очертания могут быть овальными, круглыми, эллиптическими, треугольными и другими. Листовая пластинка бывает удлиненной. Свободный конец её может быть острым, тупым, заострённым, остроконечным. Основание её сужено и оттянуто к стеблю, может быть округлым, сердцевидным.

Прикрепление листьев к стеблю
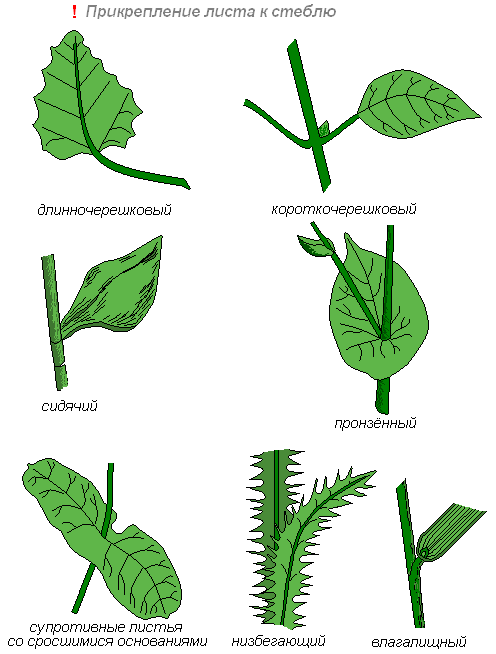
Листья прикрепляются к побегу длинными, короткими черешками или бывают сидячими.
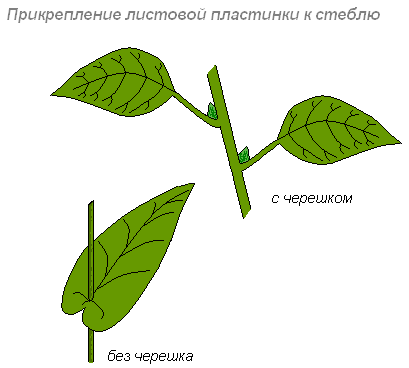
У некоторых растений основание сидячего листа на большом протяжении срастается с побегом (низбегающий лист) или побег пронизывает листовую пластинку насквозь (пронзённый лист).
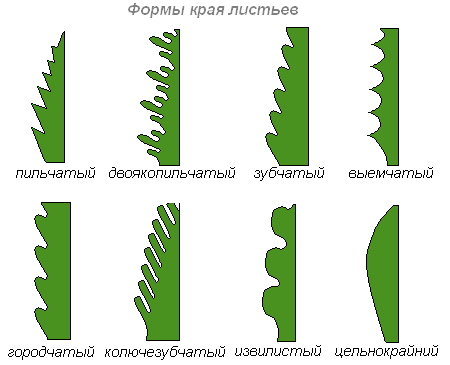
Форма края листовой пластинки
Листовые пластинки различают по степени рассечённости: неглубокие надрезы — зубчатые или пальчатые края листа, глубокие вырезы — лопастные, раздельные и рассечённые края.
Если края листовой пластинки не имеют никаких выемок, лист называется цельнокрайним . Если выемки по краю листа неглубокие, лист называется цельным .
Лопастной лист — лист, пластинка которого расчленена на лопасти до 1/3 ширины полулиста.
Раздельный лист — лист с пластинкой, расчленённой до ½ ширину полулиста.
Рассечённый лист — лист, пластинка которого расчленена до главной жилки или до основания листа.
Край листовой пластинки — пильчатый (острые углы).
Край листовой пластинки — городчатый (округлые выступы).
Край листовой пластинки — выемчатый (округлые выемки).
Жилкование
На каждом листе легко заметить многочисленные жилки, особенно отчётливые и рельефные на нижней стороне листа.
Жилки — это проводящие пучки, соединяющие лист со стеблем. Функции их — проводящая (снабжение листьев водой и минеральными солями и выведение из них продуктов ассимиляции) и механическая (жилки являются опорой для листовой паренхимы и защищают листья от разрывов). Среди разнообразия жилкования различают листовую пластинку с одной главной жилкой, от которой расходятся боковые ответвления по перистому или пальчатоперистому типу; с несколькими главными жилками, различающимися толщиной и направлением распределения по пластинке (дугонервный, параллельный типы). Между описанными типами жилкования существует много промежуточных или иных форм.
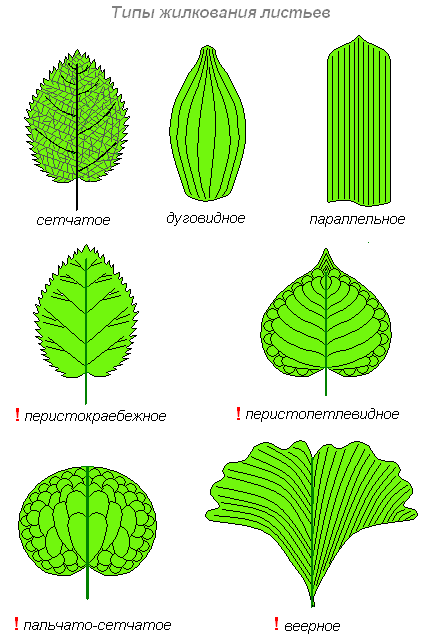
Исходная часть всех жилок листовой пластинки находится в черешке листа, откуда выходит у многих растений основная, главная жилка, разветвляясь потом в толще пластинки. По мере удаления от главной, боковые жилки всё утончаются. Самые тонкие большей частью находятся на периферии, а также вдали от периферии — посредине участков, окружённых мелкими жилками.
Существует несколько типов жилкования. У однодольных растений жилкование бывает дугонервным, при котором от стебля или влагалища вступает в пластинку ряд жилок, дугообразно направленных к вершине пластинки. У большинства злаков имеет место параллельнонервное жилкование. Дугонервное жилкование существует также у некоторых двудольных растений, например, подорожника. Однако и у них имеется связь между жилками.
У двудольных растений жилки образуют сильно разветвлённую сеть и соответственно этому различают жилкование сетчатонервоное, что говорит о лучшем обеспечении проводящими пучками.
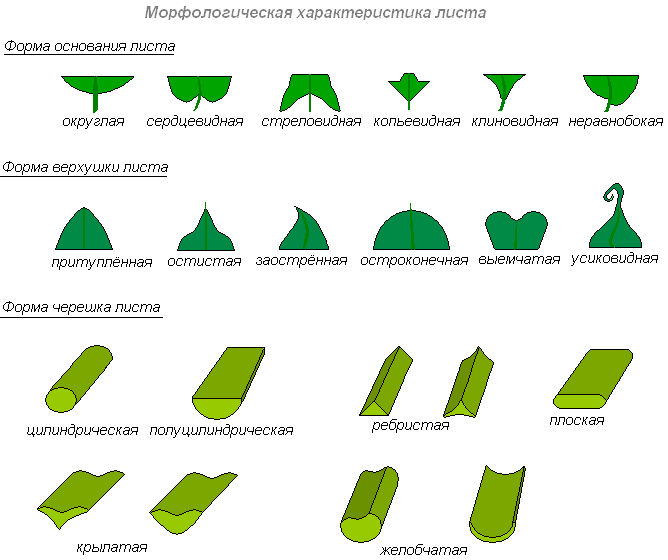
Форма основания, верхушки, черешка листа
По форме верхушки пластинки листья бывают тупые, острые, заострённые и остроконечные.
По форме основания пластинки различают листья клиновидные, сердцевидные, копьевидные, стреловидные и др.
Внутреннее строение листа
Строение кожицы листа
Верхняя кожица (эпидерма) — покровная ткань на обращённой стороне листа, часто покрытая волосками, кутикулой, воском. Снаружи лист имеет кожицу (покровную ткань), которая защищает его от неблагоприятных воздействий внешней среды: от высыхания, от механических повреждений, от проникновения к внутренним тканям болезнетворных микроорганизмов. Клетки кожицы живые, по размерам и форме они разные. Одни из них более крупные, бесцветные, прозрачные и плотно прилегают друг к другу, что повышает защитные качества покровной ткани. Прозрачность клеток позволяет проникать солнечному свету внутрь листа.
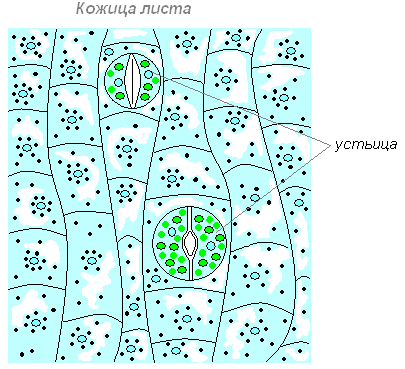
Другие клетки более мелкие, в них имеются хлоропласты, придающие им зелёный цвет. Эти клетки располагаются парами и обладают способностью изменять свою форму. При этом клетки или отдаляются друг от друга, и между ними появляется щель, или приближаются друг к другу и щель исчезает. Эти клетки назвали замыкающими, а возникающую между ними щель — устьичной. Устьице открывается, когда замыкающие клетки насыщены водой. При оттоке воды из замыкающих клеток устьице закрывается.
Строение устьица
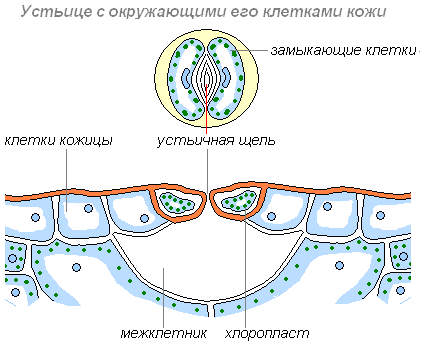
Через устьичные щели воздух поступает к внутренним клеткам листа; через них же газообразные вещества, в том числе и пары воды, выходят из листа наружу. При недостаточном обеспечение растения водой (что может случиться в сухую и жаркую погоду), устьица закрываются. Этим растения защищают себя от иссушения, так как водяные пары при закрытых устьичных щелях не выходят наружу и сохраняются в межклетниках листа. Таким образом, растения сохраняют воду в засушливый период.
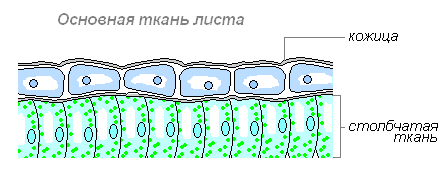
Основная ткань листа
Столбчатая ткань — основная ткань, клетки которой имеют цилиндрическую форму, плотно прилегают друг к другу и расположены с верхней стороны листа (обращённой к свету). Служит для фотосинтеза. Каждая клетка этой ткани имеет тонкую оболочку, цитоплазму, ядро, хлоропласты, вакуоль. Наличие хлоропластов придаёт зелёный цвет ткани и всему листу. Клетки, которые прилегают к верхней кожице листа, вытянуты и расположены вертикально, называют — столбчатой тканью.
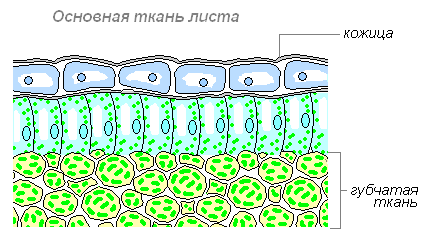
Губчатая ткань — основная ткань, клетки которой имеют округлую форму, расположены рыхло и между ними образуются крупные межклетники, также заполненные воздухом. В межклетниках основной ткани накапливаются пары воды, поступающие сюда из клеток. Служит для фотосинтеза, газообмена и транспирации (испарения).
Количество слоёв клеток столбчатой и губчатой тканей зависит от освещения. В листьях выросших на свету, столбчатая ткань развита сильнее, чем у листьев, выросших в условиях затемнения.
Проводящая ткань — основная ткань листа, пронизанная жилками. Жилки — это проводящие пучки, так как они образованы проводящими тканями — лубом и древесиной. По лубу осуществляется передача растворов сахара из листьев ко всем органам растения. Движение сахара идёт по ситовидным трубкам луба, которые образованы живыми клетками. Эти клетки вытянуты в длину, и в том месте, где они соприкасаются друг с другом короткими сторонами в оболочках, имеются небольшие отверстия. Через отверстия в оболочках раствор сахара переходит из одной клетки в другую. Ситовидные трубки приспособлены к передаче органического вещества на большое расстояние. Плотно по всей длине к боковой стенке ситовидной трубки прилегают живые клетки меньших размеров. Они сопутствуют клеткам трубки, и их называют клетками спутницами.
Строение жилок листа
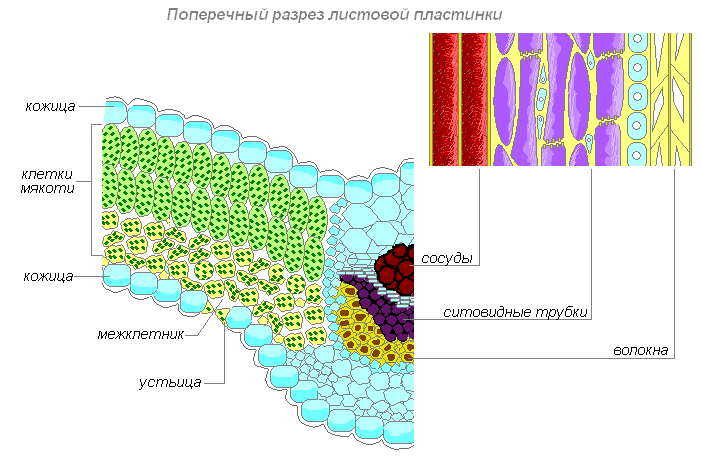
Кроме луба в состав проводящего пучка входит и древесина. По сосудам листа, так же как и в корне, движется вода с растворёнными в ней минеральными веществами. Воду и минеральные вещества растение поглощает из почвы корнями. Затем из корней по сосудам древесины эти вещества поступают в надземные органы, в том числе и к клеткам листа.
В состав многочисленных жилок входят волокна. Это длинные клетки с заострёнными концами и утолщёнными одревесневшими оболочками. Крупные жилки листа нередко окружены механической тканью, которая целиком состоит из толстостенных клеток — волокон.
Таким образом, по жилкам идёт передача раствора сахара (органического вещества) из листа к другим органам растений, а от корня — воды и минеральных веществ к листьям. Из листа растворы движутся по ситовидным трубкам, а к листу — по сосудам древесины.
Нижняя кожица покровная ткань с нижней стороны листа, обычно несёт устьица.
Жизнедеятельность листа
Зелёные листья — органы воздушного питания. Зелёный лист выполняет важную функцию в жизни растений — здесь образуются органические вещества. Строение листа хорошо соответствует этой функции: он имеет плоскую листовую пластинку, а в мякоти листа содержится огромное количество хлоропластов с зелёным хлорофиллом.
Вещества необходимые для образования крахмала в хлоропластах
Цель: выясним, какие вещества необходимы для образования крахмала?
Что делаем: поместим два небольших комнатных растения в тёмное место. Через два три дня первое растение поставим на кусок стекла, а рядом поместим стакан с раствором едкой щёлочи (она поглотит из воздуха весь углекислый газ), и всё это накроем стеклянным колпаком. Для того чтобы воздух не поступал к растению из окружающей среды, смажем края колпака вазелином.
Второе растение также поставим под колпак, но только рядом с растением поместим стакан с содой (или кусочком мрамора), смоченными раствором соляной кислоты. В результате взаимодействия соды (или мрамора) с кислотой выделяется углекислый газ. В воздухе под колпаком второго растения образуется много углекислого газа.
Оба растения поместим в одинаковые условия (на свет).
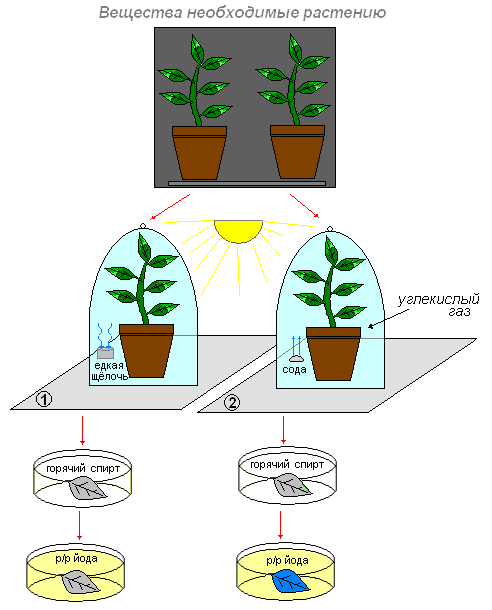
На следующий день возьмём по листу с каждого растения и обработаем вначале горячим спиртом, промываем и действуем раствором йода.
Что наблюдаем: в первом случае окраска листа не изменилась. Темно-синим стал лист того растения, которое находилось под колпаком, где был углекислый газ.
Вывод: это доказывает, что углекислый газ необходим растению для образования органического вещества (крахмал). Этот газ входит в состав атмосферного воздуха. Воздух поступает в лист через устьичные щели и заполняет пространства между клетками. Из межклетников углекислый газ проникает во все клетки.
Образование в листьях органических веществ
Цель: выяснить, в каких клетках зеленого листа образуются органические вещества (крахмал, сахар).
Что делаем: комнатное растение герань окаймлённая поместим на трое суток в тёмный шкаф (чтобы произошёл отток питательных веществ из листьев). Через трое суток вынем растение из шкафа. Прикрепим на один из листьев конверт из чёрной бумаги с вырезанным словом «свет» и поставим растение на свет или под электрическую лампочку. Через 8-10 часов срежем лист. Снимем бумагу. Опустим лист в кипящую воду, а затем на несколько минут в горячий спирт (в нём хлорофилл хорошо растворяется). Когда спирт окрасится в зелёный цвет, а лист обесцветится, промоем его водой и поместим в слабый раствор йода.
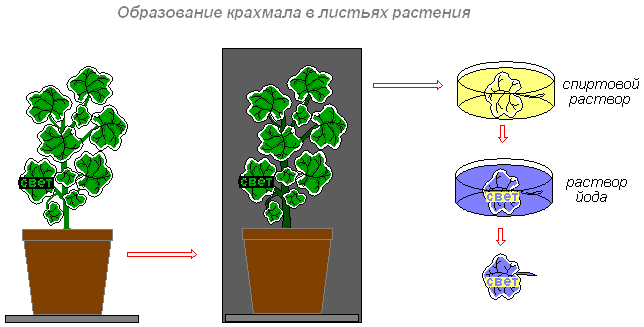
Что наблюдаем: на обесцвеченном листе появятся синие буквы (крахмал синеет от йода). Буквы появляются на той части листа, на которую падал свет. Значит, в освещённой части листа образовался крахмал. Необходимо обратить внимание на то, что белая полоска по краю листа не окрасилась. Это объясняет то, что в пластидах клеток белой полоски листа герани окаймлённой нет хлорофилла. Поэтому крахмал не обнаруживается.
Вывод: таким образом, органические вещества (крахмал, сахар) образуются только в клетках с хлоропластами, и для их образования необходим свет.
Специальные исследования учёных показали, что на свету в хлоропластах образуется сахар. Затем в результате превращений из сахара в хлоропластах образуется крахмал. Крахмал — это органическое вещество, которое в воде не растворяется.
Выделяют световую и темновую фазы фотосинтеза.
Во время световой фазы фотосинтеза происходит поглощение света пигментами, образование возбуждённых (активных) молекул, обладающих избытком энергии, идут фотохимические реакции, в которых принимают участие возбуждённые молекулы пигментов. Световые реакции протекают на мембранах хлоропласта, где находится хлорофилл. Хлорофилл является высокоактивным веществом, осуществляющим поглощение света, первичное запасание энергии и дальнейшее преобразование её в химическую энергию. В фотосинтезе принимают участие и жёлтые пигменты каротиноиды.
Процесс фотосинтеза можно представить в виде суммарного уравнения:
6СО 2 + 6Н 2 О = С 6 Н 12 О 6 + 6О 2
Таким образом, суть световых реакций заключается в том, что световая энергия превращается в химическую.
Темновые реакции фотосинтеза идут в матриксе (строме) хлоропласта при участии ферментов и продуктов световых реакций и приводят к синтезу органических веществ из углекислоты и воды. Для темновых реакций не нужно непосредственное участие света.
Итогом темновых реакций является образование органических соединений.
Процесс фотосинтеза осуществляется в хлоропластах, в два этапа. В гранах (тилакоидах) протекают реакции, вызываемые светом, — световые, а в строме — реакции, не связанные со светом, — темновые, или реакции фиксации углерода.
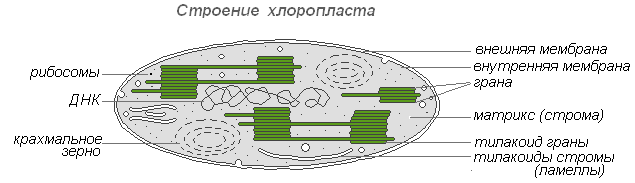
Световые реакции
1. Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбуждённое состояние. В результате этого электроны ē сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где и накапливаются, создавая отрицательно заряженное электрическое поле.
2. Место вышедших электронов в молекулах хлорофилла занимают электроны воды ē, так как вода под действием света подвергается фоторазложению (фотолизу):
Н 2 О↔ОН‾+Н + ; ОН‾−ē→ОН.
Гидроксилы ОН‾, став радикалами ОН, объединяются: 4ОН→2Н 2 О+О 2 , образуя воду и свободный кислород, который выделяется в атмосферу.
3. Протоны Н + не проникают через мембрану тилакоида и накапливаются внутри, используя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
4. При достижении критической разности потенциалов (200 мВ) протоны Н + устремляются по протонному каналу в ферменте АТФ-синтетаза, встроенному в мембрану тилакоида, наружу. На выходе из протонного канала создаётся высокий уровень энергии, которая идёт на синтез АТФ (АДФ+Ф→АТФ) . Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации углерода.
5. Протоны Н + , вышедшие на поверхность мембраны тилакоида, соединяются с электронами ē, образуя атомарный водород Н, который идёт на восстановление переносчиков НАДФ + : 2ē+2Н + =НАДФ + →НАДФ∙Н 2 (переносчик с присоединённым водородом; восстановленный переносчик).
Такими образом, активированный световой энергией электрон хлорофилла используется для присоединения водорода к переносчику. НАДФ∙Н2 переходит в строму хлоропласта, где участвует в реакциях фиксации углерода.
Реакции фиксации углерода (темновые реакции)
Осуществляется в строме хлоропласта, куда поступают АТФ, НАДФ∙Н 2 от тилакоидов гран и СО 2 из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения — пентозы С 5 , которые образуются в цикле Кальвина (цикл фиксации СО 2), Упрощённо этот цикл можно представить следующим образом:
1. К пентозе С 5 присоединяется СО 2 , в результате чего появляется нестойкое шестиугольное соединение С 6 , которое расщепляется на две трёхуглеродные группы 2С 3 — триозы.
2. Каждая из триоз 2С 3 принимает по одной фосфатной группе от двух АТФ, что обогащает молекулы энергией.
3. Каждая из триоз 2С 3 присоединяет по одному атому водорода от двух НАДФ∙Н2.
4. После чего одни триозы объединяются, образуя углеводы 2С 3 → С 6 → С 6 Н 12 О 6 (глюкоза).
5. Другие триозы объединяются, образуя пентозы 5С 3 →3С 5 , и вновь включаются в цикл фиксации СО 2 .
Суммарная реакция фотосинтеза:
6СО 2 +6Н 2 О хлорофилл энергия света →С 6 Н 12 О 6 +6О 2

Кроме углекислого газа в образовании крахмала принимает участие вода. Её растение получает из почвы. Корни поглощают воду, которая по сосудам проводящих пучков поднимается в стебель и далее в листья. А уже в клетках зелёного листа, в хлоропластах, из углекислого газа и воды при наличии света образуется органическое вещество.
Что происходит с органическими веществами, образованными в хлоропластах?
Образовавшийся в хлоропластах крахмал под воздействием особых веществ превращается в растворимый сахар, который поступает к тканям всех органов растения. В клетках некоторых тканей сахар может вновь превратиться в крахмал. Запасной крахмал накапливается в бесцветных пластидах.
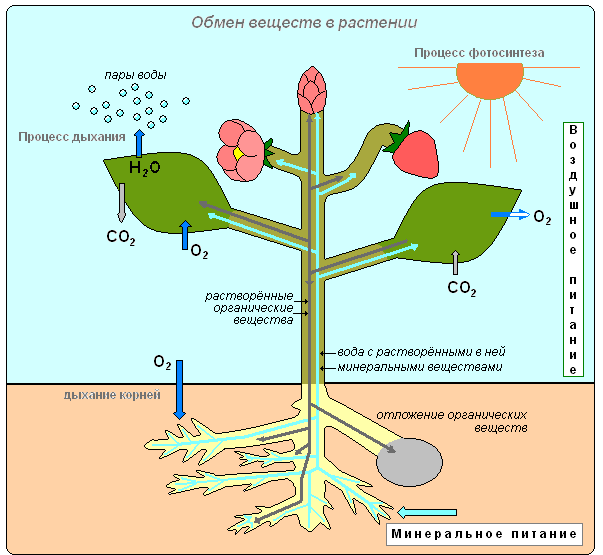
Из сахаров, образовавшихся при фотосинтезе, а также минеральных солей, поглощённых корнями из почвы, растение создаёт вещества, которые ему необходимы: белки, жиры и многие другие белки, жиры и многие другие.
Часть органических веществ, синтезированных в листьях, расходуется на рост и питание растения. Другая часть откладывается в запас. У однолетних растений запасные вещества откладываются в семенах, плодах. У двулетних на первом году жизни они накапливаются в вегетативных органах. У многолетних трав вещества запасаются в подземных органах, а у деревьев и кустарников — в сердцевине, основной ткани коры и древесины. Кроме того, у них на определённом году жизни органические вещества начинают запасаться также в плодах и семенах.
Типы питания растения (минеральное, воздушное)
В живых клетках растения постоянно происходит обмен веществ и энергии. Одни вещества поглощаются и используются растением, другие выделяются в окружающую среду. Из простых веществ образуются сложные. Сложные органические вещества расщепляются на простые. Растения накапливает энергию, а в процессе фотосинтеза и освобождает её при дыхании, используя эту энергию для осуществления различных процессов жизнедеятельности.
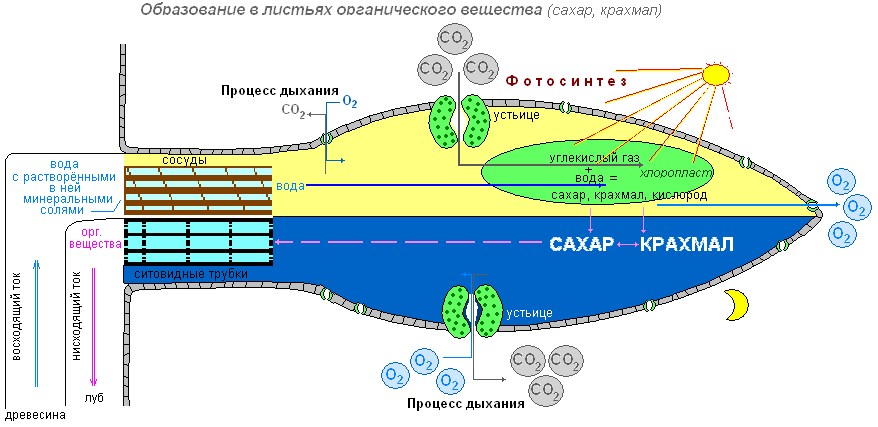
Газообмен
Листья благодаря работе устьиц осуществляют и такую важную функцию, как газообмен между растением и атмосферой. Через устьица лист с атмосферным воздухом поступают углекислый газ и кислород. Кислород используется при дыхании, углекислый газ необходим растению для образования органических веществ. Через устьица в воздух выделяется кислород, который образовался в процессе фотосинтеза. Удаляется и углекислый газ, появившийся у растения в процессе дыхания. Фотосинтез осуществляется только на свету, а дыхание на свету и в темноте, т.е. постоянно. Дыхание во всех живых клетках органов растения происходит непрерывно. Как и животные, растения погибают с прекращением дыхания.
В природе происходит обмен веществ между живым организмом и окружающей средой. Поглощение растением одних веществ из внешней среды сопровождается выделением других. Элодея, будучи водным растением, использует для питания углекислый газ, растворённый в воде.
Цель: выясним, какое же вещество выделяет элодея во внешнюю среду при фотосинтезе?
Что делаем: стебли веточек подрежем под водой (вода кипяченная) у основания и прикроем стеклянной воронкой. Пробирку, до краёв заполненную водой помещаем на трубку воронки. Это сделать в двух вариантах. Одну ёмкость поставить в тёмное место, а другую — выставить на яркий солнечный или искусственный свет.

В третью и четвёртую ёмкости добавить углекислый газ (добавить небольшое количество питьевой соды или можно подышать в трубочку) и так же один поставить в темноту другой на солнечный свет.

Что наблюдаем: через некоторое время в четвёртом варианте (сосуд, стоящий на ярком солнечном свете) начинают выделяться пузырьки. Этот газ вытесняет из пробирки воду, её уровень в пробирке вытесняется.
Что делаем: когда вода будет вытеснена газом полностью, необходимо осторожно снять пробирку с воронки. Плотно закрыть отверстие большим пальцем левой руки, а правой быстро внести в пробирку тлеющую лучинку.
Что наблюдаем: лучинка загорается ярким пламенем. Посмотрев на растения, которые поместили в темноту, увидим, что пузырьки газа из элодеи не выделяются, и пробирка осталась заполненная водой. То же самое с пробирками в первом и втором варианте.
Вывод: отсюда следует, что газ, который выделила элодея — кислород. Таким образом, растение выделяет кислород только тогда, когда есть все условия для фотосинтеза — вода, углекислый газ, свет.
Испарение воды листьями (транспирация)
Процесс испарения воды листьями у растений регулируется открыванием и закрыванием устьиц. Закрывая устьица, растение защищает себя от потери воды. Открывание и закрывание устьиц находится под влиянием факторов внешней и внутренней среды, в первую очередь температуры и интенсивности солнечного света.
Листья растений содержат много воды. Она поступает по проводящей системе от корней. Внутри листа вода продвигается по стенкам клеток и по межклетникам к устьицам, через которые уходит в виде пара (испаряется). Этот процесс легко проверить, если выполнить несложное приспособление, как показано на рисунке.

Испарение воды растением называется транспирацией. Воду испаряет поверхность листа растения, особенно интенсивно — поверхность листа. Различают транспирацию кутикулярную (испарение всей поверхностью растения) и устьичную (испарение через устьица). Биологическое значение транспирации состоит в том, что она является средством передвижения воды и различных веществ по растению (присасывающее действие), способствует поступлению углекислого газа внутрь листа, углеродному питанию растений, защищает листья от перегрева.
Интенсивность испарения воды листьями зависит от:
- биологических особенностей растений;
- условий роста (растения засушливых местностей испаряют мало воды, влажных — значительно больше; теневые растения испаряют воды меньше, чем световые; много воды растения испаряют в зной, значительно меньше — в облачную погоду);
- освещения (рассеянный свет уменьшает транспирацию на 30-40%);
- содержания воды в клетках листа;
- осмотического давления клеточного сока;
- температуры почвы, воздуха и тела растения;
- влажности воздуха и скорости ветра.
Наибольшее количество воды испаряется у некоторых видов древесных пород через листовые рубцы (рубец, оставляемый опавшими листьями на стебле), которые оказываются наиболее уязвимыми местами на дереве.
Взаимосвязь процессов дыхания и фотосинтеза
Весь процесс дыхания протекает в клетках растительного организма. Он состоит из двух этапов, в ходе которых органические вещества расщепляются на углекислый газ и воду. На первом этапе при участии специальных белков (ферментов) происходит распад молекул глюкозы на более простые органические соединения и выделяется немного энергии. Этот этап дыхательного процесса происходит в цитоплазме клеток.
На втором этапе простые органические вещества, образовавшиеся на первом этапе, под действием кислорода распадаются на углекислый газ и воду. При этом высвобождается много энергии. Второй этап дыхательного процесса протекает только с участием кислорода и в специальных тельцах клетки.
Поглощённые вещества в процессе преобразований в клетках и тканях становятся веществами, из которых растение строит своё тело. Все преобразования веществ, происходящее в организме, всегда сопровождаются потреблением энергии. Зелёное растение, как автотрофный организм, поглощая световую энергию Солнца, накапливает её в органических соединениях. В процессе дыхания при расщеплении органических веществ эта энергия высвобождается и используется растением для процессов жизнедеятельности, которые происходят в клетках.
Оба процесса — фотосинтез и дыхание — идут путём последовательных многочисленных химических реакций, в которых одни вещества преобразуются в другие.

Так, в процессе фотосинтеза из углекислого газа и воды, полученных растением из окружающей среды, образуются сахара, которые затем превращаются в крахмал, клетчатку или белки, жиры и витамины — вещества, необходимые растению для питания и запасания энергии. В процессе дыхания, наоборот, происходит расщепление созданных в процессе фотосинтеза органических веществ на неорганические соединения — углекислый газ и воду. При этом растение получает высвобождающуюся энергию. Эти превращения веществ в организме называют обменом веществ. Обмен веществ — один из важнейших признаков жизни: с прекращением обмена веществ прекращается жизнь растения.
Влияние факторов среды на строение листа
Листья растений влажных мест, как правило, крупные с большим количеством устьиц. С поверхности этих листьев испаряется много влаги.
Листья растений засушливых мест невелики по размеру и имеют приспособления, уменьшающие испарение. Это густое опушение, восковой налёт, относительно небольшое число устьиц и др. У некоторых растений листья мягкие и сочные. В них запасается вода.
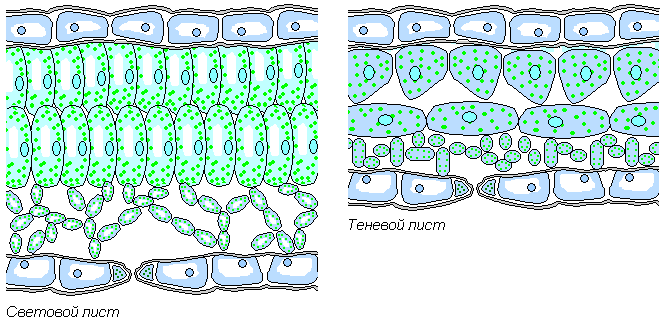
Листья теневыносливых растений имеют всего два-три слоя округлых, неплотно прилегающих друг к другу клеток. Крупные хлоропласты расположены в них так, что не затеняют друг друга. Теневые листья, как правило, более тонкие и имеют более тёмную зелёную окраску, так как содержат больше хлорофилла.
У растений открытых мест мякоть листа насчитывает несколько слоев, плотно прилегающих друг к другу столбчатых клеток. В них содержится меньше хлорофилла, поэтому световые листья имеют более светлую окраску. Те и другие листья иногда можно встретить и в кроне одного и того же дерева.
Защита от обезвоживания
Наружная стенка каждой клетки кожицы листа не только утолщена, но и защищена кутикулой, которая плохо пропускает воду. Защитные свойства кожицы значительно повышаются при образовании волосков, которые отражают солнечные лучи. Благодаря этому нагревание листа понижается. Всё это ограничивает возможность испарения воды с поверхности листа. При недостатке воды закрывается устьичная щель и пар не выходит наружу, накапливаясь в межклетниках, что приводит к прекращению испарения с поверхности листа. Растения жарких и сухих мест обитания имеют небольшую пластинку. Чем меньше поверхность листа, тем меньше опасность излишней потери воды.
Видоизменения листьев
В процессе приспособления к условиям окружающей среды листья у некоторых растений видоизменились потому, что стали играть роль не свойственную типичным листьям. У барбариса часть листьев видоизменились в колючки.

Старение листьев и листопад
Листопаду предшествует старение листьев. Это значит, что во всех клетках снижается интенсивность жизненных процессов — фотосинтеза, дыхания. Уменьшается содержание уже имеющихся в клетках важных для растения веществ и сокращается поступление новых, в том числе и воды. Распад веществ преобладает над их образованием. В клетках накапливаются ненужные, и даже вредные продукты, их называют конечными продуктами обмена веществ. Эти вещества удаляются из растения при сбрасывании листьев. Наиболее же ценные соединения по проводящим тканям оттекают из листьев в другие органы растения, где откладываются в клетках запасающих тканей или сразу используется организмом для питания.
У большинства деревьев и кустарников в период старения листья меняют окраску и становятся жёлтыми или багряными. Это происходит потому, что хлорофилл разрушается. Но помимо него в пластидах (хлоропластах) имеются вещества желтого и оранжевого цвета. Летом они были, как бы замаскированы хлорофиллом и пластиды имели зелёный цвет. Кроме того, в вакуолях накапливаются другие красящие вещества жёлтого или красно-малинового цвета. Вместе с пигментами пластид они определяют окраску осенних листьев. У некоторых растений листья сохраняют зелёный цвет до отмирания.
Ещё до того как с побега упадёт лист, в его основании на границе со стеблем формируется слой пробки. Наружу от него образуется отделительный слой. Со временем клетки этого слоя оделяются друг от друга, так как ослизняется и разрушается межклеточное вещество, которое их соединяло, а иногда и оболочки клеток. Лист отделяется от стебля. Однако некоторое время он ещё сохраняется на побеге благодаря проводящим пучкам между листом и стеблем. Но наступает момент нарушения и этой связи. Рубец на месте отделившегося листа покрыт защитной тканью, пробкой.
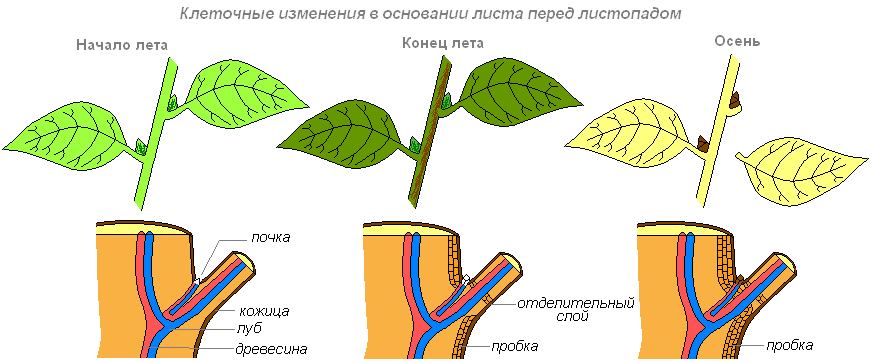
Как только листья достигают предельных размеров, начинаются процессы старения, ведущие, в конце концов, к отмиранию листа — его пожелтение или покраснение, связанное с разрушением хлорофилла, накоплением каротиноидов и антоцианов. По мере старения листа снижается также интенсивность фотосинтеза и дыхания, деградируют хлоропласты, накапливаются некоторые соли (кристаллы оксалаты кальция), из листа оттекают пластические вещества (углеводы, аминокислоты).
В процессе старения листа близ его основания у двудольных древесных растений формируется так называемый отделительный слой, который состоит из легко расслаивающейся паренхимы. По этому слою лист и отделяется от стебля, причём на поверхности будущего листового рубца заранее образуется защитный слой пробковой ткани.

На листовом рубце заметны в виде точек поперечные сечения листового следа. Скульптура листового рубца различна и является характерным признаком для систематики лепидофитов.
У однодольных и травянистых двудольных отделительный слой, как правило, не образуется, лист отмирает и разрушается постепенно, оставаясь на стебле.
У листопадных растений опадение листьев на зиму имеет приспособительное значение: сбрасывая листья, растения резко уменьшают испаряющую поверхность, защищаются от возможных поломок под тяжестью снега. У вечнозелёных растений массовый листопад приурочен обычно к началу роста новых побегов из почек и поэтому происходит не осенью, а весной.
Осенний листопад в лесу имеет важное биологическое значение. Опавшие листья — хорошее органическое и минеральное удобрение. Ежегодно в на их лиственных лесах опавшие листья служат материалом для минерализации, производимой почвенными бактериями и грибами. Кроме того, опавшая листва стратифицирует семена, опавшие до листопада, предохраняет корни от вымерзания, препятствует развитию мохового покрова и т.д. некоторые виды деревьев сбрасывают не только листву, но и годовалые побеги.
Устьица растений
находятся в их кожице (эпидермисе). Каждое растение находится в постоянном обмене с окружающей атмосферой. Оно постоянно поглощает кислород и выделяет углекислоту. Кроме того, своими зелеными частями оно поглощает углекислоту и выделяет кислород. Затем, растение постоянно испаряет воду. Так как кутикула, которой покрыты листья и молодые стебли, очень слабо пропускает через себя газы и водяные пары, то для беспрепятственного обмена с окружающей атмосферой в кожице имеются особые отверстия, называемые У. На поперечном разрезе листа (фиг. 1) У. представляется в виде щели (S
), ведущей в воздухоносную полость (i
).
Фиг. 1. Устьице (S
) листа гиацинта в разрезе. По обеим сторонам У. находится по одной замыкающей клетке.
Оболочки замыкающих клеток дают в сторону устьичного отверстия по два выроста, благодаря которым оно распадается на две камеры: передний и задний дворик. При рассматривании с поверхности - У. представляется в виде продолговатой щели, окруженной двумя полулунными замыкающими клетками (фиг. 2). Днем У. открыты, на ночь же они закрываются. У. закрываются также днем во время засухи. Закрывание У. производится замыкающими клетками. Если кусочек кожицы листа положить в воду, то У. продолжают оставаться открытыми. Если же воду заменить раствором сахара, вызывающим плазмолиз клеток, то У. закроются. Так как плазмолиз клеток сопровождается уменьшением их объема, то отсюда следует, что закрывание У. есть результат уменьшения объема замыкающих клеток.
Во время засухи замыкающие клетки теряют часть своей воды, уменьшаются в объеме и закрывают У. Лист оказывается покрытым сплошным слоем кутикулы, слабо пропускающей водяные пары, чем и предохраняется от дальнейшего высыхания. Ночное закрывание У. объясняется следующими соображениями. Замыкающие клетки постоянно содержат в себе хлорофилловые зерна и поэтому способны к усвоению атмосферной углекислоты, т. е. к самостоятельному питанию. Накопленные на свету органические вещества сильно притягивают к себе из окружающих клеток воду, поэтому замыкающие клетки увеличиваются в объеме и открываются. Ночью же выработанные на свету органические вещества расходуются, а вместе с ними утрачивается способность притягивать воду, и У. закрываются. У. находятся как на листьях, так и на стеблях. На листьях они помещаются или на обеих поверхностях, или же на одной из них. Травянистые, мягкие листья имеют У. как на верхней, так и на нижней поверхности. Твердые кожистые листья имеют У. почти исключительно на нижней поверхности. У листьев, плавающих на поверхности воды, У. исключительно находятся на верхней стороне. Количество У. у различных растений очень различно. Для большинства листьев число У., находящихся на одном квадратном миллиметре, колеблется между 40 и 300. Наибольшее число У. находится на нижней поверхности листа Brassica Rapa - на 1 кв. мм 716. Существует некоторая зависимость между количеством У. и влажностью места. В общем растения влажных местностей имеют более У., чем растения сухих местностей. Кроме обыкновенных У., служащих для газового обмена, у многих растений имеются еще водяные
У. Они служат для выделения воды не в газообразном состоянии, но в жидком. Вместо лежащей под обыкновенными У. воздухоносной полости под водяными У. находится особая водоносная ткань, состоящая из клеток с тонкими оболочками. Водяные У. встречаются по большей части у растений сырых местностей и находятся на различных частях листьев независимо от находящихся тут же обыкновенных У. Водяные У. выделяют капли воды по большей части тогда, когда вследствие большой влажности воздуха не могут испарять воду воздухоносные У. Кроме водяных У., существует целый ряд разнообразных приспособлений для выделения листьями воды в жидком виде. Все такие образования получили название гидатод
(Hydathode). Примером могут служить гидатоды Gonocaryum pyriforme (фиг. 3). На поперечном разрезе через лист видно, что некоторые из клеток кожицы особенным образом изменились и превратились в гидатоды. Каждая гидатода состоит из трех частей. Наружу выдается идущий вкось вырост, пронизанный узким канальцем, через который вытекает вода гидатоды. Средняя часть имеет вид воронки с очень утолщенными стенками. Нижняя часть гидатоды состоит из тонкостенного пузыря. Некоторые растения выделяют своими листьями большие количества воды, не имея никаких особо устроенных гидатод. Напр. различные виды Salacia выделяют между 6-7 часами утра такие большие количества воды, что вполне заслуживают названия дождевых кустарников: при легком прикосновении к ветвям с них падает настоящий дождь. Вода выделяется простыми порами, покрывающими в большом количестве наружные оболочки клеток кожицы. В. Палладин.


Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона. - С.-Пб.: Брокгауз-Ефрон . 1890-1907 .
Смотреть что такое "Устьица растений" в других словарях:
Находятся в их кожице (эпидермисе). Каждое растение находится в постоянном обмене с окружающей атмосферой. Оно постоянно поглощает кислород и выделяет углекислоту. Кроме того, своими зелеными частями оно поглощает углекислоту и выделяет кислород …
Устьице листа томата под электронным микроскопом Устьице (лат. stoma, от греч. στόμα «рот, уста») в ботанике это пора, находящаяся на нижнем или верхнем слое эпидермиса листа растения, через которое происходит испарение воды и газообмен с… … Википедия
Первые попытки классификации цветковых растений, как и растительного мира вообще, были основаны на немногих, произвольно взятых, легко бросающихся в глаза внешних признаках. Это были чисто искусственные классификации, в которых в одной… … Биологическая энциклопедия
Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Группы клеток, расположенные в теле растения известным порядком, имеющие определенное строение и служащие для различных жизненных отправлений растительного организма. Клетки почти всех многоклеточных растений не однородны, а собраны в Т. У низших … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона - представляют из себя такого рода процессы и явления, происходящие в живом растительном организме, которые при нормальной его жизнедеятельности никогда не встречаются. По определению Франка, Б. растений это отклонение от нормального состояния вида … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Содержание: Предмет Ф. Ф. питания. Ф. роста. Ф. формы растений. Ф. размножения. Литература. Ф. растения изучает процессы, совершающиеся в растениях. Эта часть обширной науки о растениях ботаники отличается от ее остальных частей систематики,… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Лист (folium), орган высших растений, выполняющий функции фотосинтеза и транспирации, а также обеспечивающий газообмен с воздушной средой и участвующий в др. важнейших процессах жизнедеятельности растения. Морфология, анатомия листа и его… … Большая советская энциклопедия
ПОКРОВНЫЕ ТКАНИ.
- Общая характеристика и св-ва.
- Строение и ф-ции первичной покровной ткани – эпидермы.
Функции эпидермы
Осн. клетки эпидермы
- Строение вторичной покровной ткани – перидермы
Образование и заложение
Строение чечевичек
- Строение корки – ретидома.
Общая характеристика и св-ва.
Покровные ткани покрывают тело растения сверху, т.е. располагаются на поверхности и непосредственно контактируют с внешней средой. С одной стороны, они защищают растения от всяких неблагоприятных воздействий (от проникновения микроорганизмов, от испарения влаги), поэтому клетки покровных тканей плотно сомкнуты, без межклетников. С другой стороны, они должны обеспечивать связь растений с внешней средой (газообмен, транспирацию), поэтому в них имеются особые структуры.
Таким образом, строение и свойства покровных тканей определяются выполняемыми ими функциями.
Свойства: 1. Многофункциональность, т.е. это сложные ткани, состоят из нескольких типов клеток, выполняющих разные функции.
2. Обеспечивают избирательная проницаемость (для газов, Н 2 О и др. в-в), для чего имеются специальные структуры.
3. Постоянная смена тканей с возрастом, или изменение их функций.
Различают: первичную, вторичную и третичную покровные ткани, которые сменяют друг друга с возрастом.
2. Первичная покровная ткань – эпидерма (кожица).
Возникла как результат эволюционного приспособления растений к наземным условиям жизни.
(Эпи – греч. «сверху», «над», дерма – «кожа»).
а) Возникает из наружного слоя апикальной (верхушечной) меристемы побега – протодермы, следовательно – это первичная покровная ткань; защищает и покрывает листья, молодые стебли, цветки и плоды высших растений.
Эпидерма – первичная покровная ткань растения.
Обычно однослойная, состоит из 3 типов клеток, выполняющих разные ф-ции, т.е. это сложная ткань.
Включает:
1) плотно сомкнутые основные клетки эпидермы,
2) замыкающих и побочные клетки устьиц,
3) трихомы – производные эпидермы в виде выростов и волосков.
Функции и свойства эпидермы определяются особенностями строения этих клеток, которые идеально приспособлены для наземного существования растений.
(При переходе к вторично водному образу жизни цветковые растения, например, могут утратить устьица и защитные структуры основных клеток эпидермы).
1. Основные клетки эпидермы .
1) Образуют 1 слой живых, прозрачных клеток (т.к. располагаются на растущих органах), следовательно хорошо пропускают свет к расположенным ниже ассим. клеткам, где идет фотосинтез.
Защитную и покровную функцию обеспечивают другие особенности:
2) плотно сомкнуты, без межклетников
3) толстые внешние стенки клеток, тонкие боковые
4) боковые стенки клеток часто имеют волнистую форму, что улучшает их смыкание и прилегание друг к другу.
5) Оболочка клетки устроена сложно: нижняя часть состоит из целлюлозы, а верхняя кутинизирована.
6) Очень характерно образование на поверхности оболочки слоя кутикулы, регулирующей уменьшение испарения и газообмен.
Кутикула – тонкий, прозрачный слой кутина и растительных восков на наружных стенках основных клеток. Воск – жироподобное в-во, уменьшает проницаемость для воды и газов. Следовательно, мощность кутикулы, распределение восков и кутина, число и характер пор определяют проницаемость для растворов и газов, химическую стойкость, бактерицидную устойчивость.
Вовлажном состоянии кутикула более проницаема для газов и жидкостей, чем в сухом, поэтому в дождь через кутикулу идет поглощение водных растворов (внекорневая подкормка растений).
7) Хорошо развиты ап. Гольджи и ЭПС, следовательно в осн. клетках эпидермы идет синтез ряда в-в (углеводов, белков и т.д.) (биосинтетическая функция эпидермы)
Замыкающие и побочные клетки устьиц.
Устьица, или устьичный аппарат – это две замыкающих клетки бобовидной формы с разноутолщенными стенками и с расположенным между ними межклетником - устьичной щелью (внутренние стенки устьичных клеток, обращённые к щели толстые, а наружные тонкие).
Реже рядом с ними находятся побочные клетки эпидермы, отличающиеся от ее основных клеток.
Под устьицем расположена подустьичная воздушная полость, окруженная клетками паренхимы и связанная с системой межклетников органа.
Характерно:
1) Есть хлоропласты (фотосинтез углеводов) и митохондрии (синтез АТФ). Необходимы, т.к. раскрытие устьичной щели происходит активно с затратой энергии АТФ, а смыкание пассивно.
2) За счёт разноутолщенности стенок замыкающих клеток образуется щель.
Следовательно, осн. функции :
1 - транспирация (регулируемое испарение воды)
Газообмен.
(Самостоятельно, изучить механизм работы устьиц).
Механизм открывания и закрывания устьичной щели основан на осмотических явлениях, а также принимают участие все органоиды протопласта замыкающих клеток.
- - K-Na – насос (механизм быстрого реагирования). При высоком содержании в растении воды в плазмалемме замыкающих клеток работают белки-переносчики К + , активно, с затратой энергии АТФ перекачивающие ионы К + из окружающих клеток в цитоплазму, и далее они поступают в вакуоли замыкающих клеток устьиц. При дальнейшем повышении концентрации ионов К + , вслед за ними, пассивно, по градиенту концентрации, в замыкающие клетки устьиц из окружающих клеток поступает вода. Объём вакуолей увеличивается, осмотическое давление в замыкающих клетках повышается, оболочки клеток растягиваются. Из-за того, что оболочки замыкающих клеток утолщены неравномерно, наружная оболочки растягиваются сильнее, а внутренние (обращённые друг к другу) не могут растянуться и расходятся, между ними образуется щель – устьица открываются. При недостатке воды в растении белки переносчики ионов К + прекращают свою работу. К-Nа – насос останавливается. При этом ионы К + по градиенту концентраций пассивно покидают устьичные клетки и уходят в окружающие клетки, вслед за ними уходит вода. Замыкающие клетки теряют тургор и пассивно смыкаются – устьица закрываются.
Т.о., открытие устьичной щели происходит активно, с затратой энергии, источником которой являются углеводы, накапливаемые хлоропластами. Закрытие щели происходит пассивно, без затрат энергии.
- – Суточное изменение концентрации сахаров (днем много, к концу ночи – мало) (т.к. есть хлоропласты). В течение дня в хлоропластах замыкающих клеток идёт фотосинтез. Следовательно, на свету в них повышается концентрации глюкозы и по мере её накопления, к вечеру, по градиенту концентрации внутрь клеток поступает вода. Замыкающие клетки приобретают упругое тургорное состояние, устьица раскрываются. Ночью фотосинтез прекращается, глюкоза расходуется на дыхание замыкающих клеток, её концентрация снижается. Из-за этого по градиенту концентрации вода покидает замыкающие клетки, они теряют тургор и спадаются. Устьичная щель закрывается. Утром процесс повторяется. Сейчас считается, что этот механизм ограничен, основным является работа К-Nа – насоса.
Разные растения выработали определённый ритм работы устьичного аппарата. У большинства растений устьица открыты и днём и ночью, закрываясь лишь при уменьшении содержания воды. В жаркие часы устьица обычно закрыты, но у растений пустынь они открыты (с помощью испарения они охлаждают поверхность листьев и активнее поглощают воду). У некоторых растений, например у каланхоэ, устьица открываются ночью и закрываются днём.
Типы устьиц:
Важны при установлении родства таксонов растений.
1.- анамоцитные устьица – имеют побочные клетки, не отличающиеся от осн. клеток эпидермы (хар-ны для всех групп растений, кроме хвощей).
2.- диацитные устьица – имеют две побочные клетки, общая стенка которых перпендикулярна устьичной щели (есть у ряда цветковых, у сем. Губоцветных и Гвоздичных).
3. – парацитные устьица - побочные клетки располагаются параллельно замыкающим клеткам и устьичной щели (есть папоротников, хвощей и ряда цветковых растений).
4. – анизоцитные устьица – замыкающие клетки окружены тремя побочными, одна из которых или крупнее, или меньше остальных (только у цветковых растений).
5. – тетрацитные устьица - имеют 4 побочные клетки (есть у однодольных).
6. – энциклоцитные устьица – побочные клетки образуют узкое кольцо вокруг замыкающих клеток (есть у папоротников, голосеменных и ряда цветковых).
Если устьица открыты, испарение идет так, будто эпидермы нет совсем.
Число и расположение устьиц варьирует у разных растений. В среднем, у растения 100-700 устьиц на 1 мм 2 . Обычно, у наземных растений большая часть устьиц расположена на нижней стороне листьев, сверху их очень мало или нет совсем. Это объясняется следующими причинами: 1) сверху листья сильнее нагреваются, следовательно, испарение будет идти интенсивнее и потери влаги будут больше; 2) углекислый газ образуется в почве и поднимается вверх, попадая сразу в устьица, к тому же, он тяжелее воздуха и скапливается обычно в нижних воздушных слоях.
У растений с вертикально стоящими листьями (ребром), например, у эвкалипта, устьица распределены по обеим сторонам листа. У водных растений с плавающими листьями устьица расположены на верхней стороне.
- Трихомы – волоски, выросты и производные эпидермы.
Форма, строение – систематический признак для определения видов и родов растений.
Бывают: железистые и кроющие.
Железистые трихомы образуют и накапливают экскреты и секреты. Это эфирные масла (герань, мята и др.) или защитные в-ва (крапива). Капля масла выделяется под кутикулу, накапливается там, затем при разрыве кутикулы выходит наружу. Кутикула восстанавливается и под ней накапливается новая капля эфирного масла.
Функции:
1-защитная
2-терморегуляция
3- бактерицидная (эфирные масла – фитонциды)
Кроющие трихомы состоят:
1- часто из мертвых клеток заполненных воздухом (одно- или многоклеточные мертвые трихомы)
Функции: отражение солнечных лучей, благоприятные микроклиматические условия для работы устьиц, уменьшение нагрева. Следовательно, снижение транспирации и мех. защита (растения пустынь и степей).
2- редко из живых клеток.
Функции: увеличивают общую испар. поверхность. Следовательно, увеличение испарения (растения влажных тропиков и тропических болот).
Т.о., эпидерма как многофункциональная сложная ткань выполняет функции только в живом виде.
1. Регуляция газообмена и транспирации (между растением и окр. средой)
2. Защитная (мех., бактерицидная, от излишнего испарения воды).
3. Выделительная (выделяет соли, воду, эфирные масла).
4. Поглотительная (всасывающая) (вода с мин. в-вами----внекорневые подкормки).
5. Биосинтетическая (синтез орг. в-в: кутинов, восков и т.п.)
Практич. исп.
- эпидермальные волоски (хлопчатник --- ткань).
- эфирные масла --- парфюмерия и медицина.
При дальнейшем росте стебля на 1 место выдвигается обеспечение мех. прочности, защита от потери воды и резких перепадов температур, поэтому эпидерма заменяется вторичной покровной тканью – перидермой.
3. Вторичная покровная ткань – перидерма.
П. – многослойная, вторичная покровная ткань.
Возникает после эпидермы. Ее образование начинается с закладки вторичной меристемы – феллогена (пробкового камбия). Он закладывается в однолетних побегах деревьев и кустарников к середине лета.
Его образование может идти 3 способами:
1) чаще всего закладывается в клетках субэпидермального слоя (под эпидермой)
2) иногда феллоген образуется в более глубоких слоях коры (малина, смородина),
3) редко феллоген обр-ся в самой эпидерме при тангенциальном делении ее клеток.
(верхняя клетка остается эпидермальной, а нижняя становится клеткой феллогена).
Кроме феллогена в состав перидермы входят еще 2 типа клеток, следовательно это тоже сложная ткань.
Типы клеток перидермы:
1) феллема (пробка) --- защитная функция
2) феллоген (пробковый камбий) --- образовательная функция
3) феллодерма --- выполняет питательную функцию по отношению к феллогену.
Строение перидермы.
1. Основа – феллоген , однослойная меристема, возникающая из живых паренхимных клеток постоянных тканей, следовательно – вторичная тк.
Его клетки делятся тангентально и откладывают кнаружи много слоев клеток пробки – феллемы (главные клетки), а внутрь клетки феллодермы .
Сначала клетки пробки тонкостенные, затем они опробковевают (на внутренней поверхности клеточных оболочек откладывается изолятор – суберин) и живое содержимое их отмирает и заполняется воздухом (следовательно, такие мертвые клетки приобретают хорошие изолирующие свойства).
В таком состоянии они выполняют основные функции:
1) защитная (механическая, защита от потери влаги, бактерицидная защита, т.к. устойчива к разрушению);
2) терморегуляционная (пробка терморегулятор, т.к. хорошо защищает растение от перепадов t o , следовательно, откладывается у древесных растений к концу лета – началу осени (благодаря пробке растение постепенно промерзает и постепенно оттаивает, что не дает образоваться в цитоплазме клеток кристаллов льда, которые повреждают и разрушают клетку).
Феллоген залечивает раны при повреждении растения и при благоприятных условиях постоянно формирует новые слои пробки. Поэтому перидерма – многослойная ткань. После развития перидермы, эпидерма, отрезанная от притока питательных веществ и Н 2 О, отмирает и слущивается (зеленая окраска стебля заменяется на коричневатую).
Лежащие под пробкой-феллемой живые ткани нуждаются в газообмене, поэтому в перидерме формируются чечевички – проходные разрывы в пробке, через которые происходит газообмен. Обычно чечевички закладываются под устьицами. Вместо пробки феллоген здесь откладывает живые клетки паренхимы.
Поперечный срез чечевички показывает, что слои пробки, чередуются с паренхимными клетками с хорошо развитыми межклетниками. По межклетникам и идет газообмен. Феллоген подстилает паренхимную ткань, т.к. в нем есть межклетники, он не препятствует газообмену.
К зиме феллоген откладывает под паренхимным слоем клеток замыкающий слой мертвых клеток пробки-феллемы – чечевичка закрывается (т.к. зимой растение резко сокращает газообмен и обменные процессы почти до 0).
Весной под напором новых паренхимных клеток, откладываемых феллогеном, этот слой разрывается и газообмен восстанавливается.
Чечевички у разных видов деревьев и кустарников имеют свою форму и размер и изменяются с возрастом.
Часто у деревьев на смену перидерме приходит корка – ретидом – третичная покровная ткань(например, у сосны, яблони, ясеня, клена; у платана и эвкалипта – корки нет).
Корка очень мощная, толстая, трещиноватая.
Образование .
Корка образуется при многократном заложении слоев феллогена, а затем и перидермы в глубоких тканях коры.
Живые клетки, оказавшись между этими слоями, погибают и становятся частью корки.
Т.о., корка состоит из чередующихся слоев пробки и клеток других, отмерших тканей коры, т.е. – сложная по составу.
Из-за того, что мертвые ткани пробки не могут растягиваться при росте стебля в толщину на ретидоме образуются трещины на мертвых участках.
1) защитная (увеличивают механическую прочность, устойчивость к пожарам и т.д.);
2) терморегуляционная (предохраняет растение от резких перепадов t o).
МЕХАНИЧЕСКИЕ ТКАНИ
1. Определение
2. Осн. типы мех. тканей и их ф-ции.
3. Значение для человека.
4. Распределение мех. тканей в теле растений.
Механические ткани
а) колленхима - живая (уголковая, пластинчатая, рыхлая)
б) склеренхима – мертвая (волокна, склереиды)
1. Любое растение нуждается в опоре для сохранения своей целостности. У растений четко прослеживается эволюция механических приспособлений при адаптации к жизни в разных средах. У первичноводных водорослей, обитавших в плотной и инертной воде мех. прочность достигается благодаря клеточным целлюлозным оболочкам (т.е. только на клеточном уровне) . При выходе на сушу в разреженную среду этого становится недостаточно и у мелких наземных растений устойчивость достигается благодаря клеточным оболочкам и тургору клеток (клеточный уровень) . При увеличении линейных размеров прочности клеточных оболочек и тургора становится недостаточно , и возникают особые механические ткани, обеспечивающих опору растению (тканевый уровень) . В дальнейшем при адаптивной эволюции растения все лучше приспосабливаются к воздушно-почвенной среде и мех. ткани по-разному и более оптимально располагаются в разных органах (т.е. прочность обеспечивается на органном и организменном уровне организации).
Т.о., механические ткани – играют роль опорной системы и придают прочность растениям.
Основные функции:
1) опорная (образуют внутренний скелет органов растения)
2) защитная (обеспечивают мех. прочность и устойчивость к мех. воздействиям)
Выполняют функции в сочетании с другими тканями, следовательно играют роль арматуры и часто называются арматурными .
Механические (арматурные) ткани – специализированные ткани, в сочетании с другими образующие арматуру органов и всего тела растения.
Выделяют 2 основных типа мех. тканей: колленхиму и склеренхиму.
1. Колленхима (“colla”– клей) - состоит из живых прозенхимных клеток с тупыми и скошенными концами.
Характерно:
1. Неравномерно утолщенные оболочки
2. Нет границы между первичной и вторичной оболочками, и они не одревесневают.
3. Это живые клетки, имеют хлоропласты.
Следовательно,
4. Могут вести фотосинтез (поэтому колленхима с хлоропластами часто называется
хлоренхима и располагается непосредственно сразу под эпидермой).
5. Выполняют свои функции только в состоянии тургора. Если её клетки теряют воду, то растения увядают.
Образуются в молодых побегах, в растущих листьях непосредственно из апикальных меристем , следовательно, если бы клетки колленхимы были мертвыми, с вторичными равномерными утолщениями и одревеснением, то они не могли бы растягиваться и расти путем растяжения вслед за другими тканями, следовательно не выполняли бы свои функции. Поэтому колленхима – ткань молодых растений.
Виды колленхимы.
Уголковая к. – утолщенные по углам части оболочек 3-5 клеток сливаются и образуют 3-5-угольники;
Рыхлая к. – с межклетниками между слившимися утолщенными участками оболочек (у водных растений);
Пластинчатая к. – утолщенные части оболочек расположены параллельными слоями.
Колленхима развита в основном у двудольных растений .
Склеренхима (греч. “skleros”- твердый) – разновидность мех. ткани, обеспечивающей прочность органов и всего тела растения. Встречается у всех цветковых (и у однодольных, и у двудольных).
Характерно:
1) Клетки часто с равномерно утолщенными вторичными оболочками, которые чаще всего одревесневают (имеют большую прочность к раздавливанию, но выше хрупкость).
2) Мертвые клетки. Протопласт, как правило, отмирает после формирования оболочек.
Т.е. основные функции выполняют мертвые клетки .
Склеренхима представлена волокнами и склереидами.
Волокна – длинные прозенхимные, чаще всего одревесневшие клетки, заостренные на концах с толстыми стенками и узкой полостью. Пор мало.
(исключения, неодревесневшие лубяные волокна ряда растений, например, льна).
Волокна бывают: 1) лубяные (входят в состав луба-флоэмы) (более длинные и тонкие) и 2) древесинные (волокна либриформа) (входят в состав древесины).
Могут быть и другие волокна, которые входят в состав других тканей и располагаться группами или одиночно.
Склереиды – клетки склеренхимы, не обладающие формой волокон.
1. – округлые – каменистые клетки – брахисклереиды.
2. – ветвистые – астросклереиды и т.д.
Как и волокна образуют либо сплошные группы, слои (в скорлупе орехов, косточке вишни, сливы, персика, абрикоса), либо располагаются поодиночке или группами из нескольких клеток – идиобластов (напр., идиобласты в мякоти плодов груши, айвы).
Идиобласты (греч. idios – особый, blastos – зародыш) – клетки тканей, расположенные одиночно среди массивов других тканей (характерны для механических и выделительных тканей).
Значение для человека.
Прежде всего, волокна
1) Древесинные волокна, образуют древесину --- стройматериалы, бумага и т.п.
2) Лубяные волокна – луб (лапти, дранка и т.п.)
3) Неодревесневшие лубяные волокна пластичны ---- ткани. Лён--- льняная ткань, крапива ---посконь и т.п.
Образование мех. тканей.
Колленхима – первичная ткань, – чаще всего из апикальной основной меристемы.


